 |
| トップページ > PYP タグと発蛍光プローブを利用した生細胞タンパク質イメージング技術の開発 |
 |
||||||||||||||||||||||||||
PYP タグと発蛍光プローブを利用した生細胞タンパク質イメージング技術の開発
|
||||||||||||||||||||||||||
 |
堀 雄一郎 大阪大学大学院工学研究科・大阪大学免 疫学フロンティア研究センター、助教 JST さきがけ研究者 |
 |
菊地 和也 大阪大学大学院工学研究科・大阪大学免 疫学フロンティア研究センター、教授 |
Fluorescence protein labeling by synthetic probes is a powerful approach to investigate protein function and localization inside living cells. This chemistry-based technique utilizes a pair of a protein tag and its specific ligands connected to fluorophores. Its potential advantages are that various fluorescent molecules are available as labeling reagents, and the timing of protein labeling is easily controlled. Because of these characteristics, this method is attracting attention as an alternative to fluorescent proteins. On the other hand, in this labeling system, there is a problem that the fluorescence of free probes inside cells prevents the identification of labeled proteins. As a solution to this problem, we developed a protein labeling system based on fluorogenic probes and photoactive yellow protein (PYP) tag. In this review, we describe about the design strategy of the probes, the advances in fluorogenic protein labeling systems and its biological application.
1. はじめに
タンパク質の細胞内局在は、タンパク質の生理的役割を明らかにするうえで重要な情報となる。生細胞における局在情報を可視化するための研究ツールのうち最も汎用されているものは、2008 年のノーベル賞受賞対象となった蛍光タンパク質といえる。これまでに、蛍光波長やストークスシフトの異なる様々な蛍光タンパク質が報告されてきた 1) 。その応用範囲は、局在解析のみにとどまらず、酵素活性や金属イオンの検出系、タンパク質間相互作用解析など多岐にわたり 2) 、その有用性は多くの研究者に認められている。
一方、蛍光タンパク質は万能ではなく、いまだに解決されていない問題が存在する。例えば、蛍光タンパク質は 27kDa と比較的大きく、標的タンパク質に与える立体的影響が懸念されている。また、長波長領域において、十分な明るさの蛍光を発するタンパク質がほとんどないために、厚みのある組織や小動物におけるイメージング実験では、感度の低下が問題となることがある。
近年、これらの問題を解決するアプローチとして、合成蛍光プローブを利用した新しいタンパク質標識技術が化学者により開発された。本稿では、この化学に基づいた標識技術の原理と利点及び従来法の問題点について述べ、筆者らのタンパク質標識技術の開発経緯及び応用展開について紹介する。
2. 合成蛍光プローブとタグタンパク質を利用したタンパク質標識技術
合成蛍光プローブを用いたタンパク質標識技術のうち、特定のリガンドと特異的に結合するタンパク質をタグとして用いる方法が、注目を集めている。この方法では、リガンド部分に蛍光色素を繋いだ分子をプローブとして設計し、有機合成する。一方、タグタンパク質には標的タンパク質を遺伝子工学により融合させ、細胞内で発現させる。そのタグタンパク質をプローブにより特異的に標識することで、標的タンパク質の局在をイメージングすることができる。これまでに報告されている代表的なタグタンパク質として、HaloTag 3) 、SNAP-tag 4) 、tetracysteine tag 5) があげられ、現在市販化されている。
この標識技術の利点として、次の 3 点を挙げることができる。
1 点目は、特定のタイミングで標的タンパク質を容易に標識できることである。このためある時間に発現しているタンパク質の局在や動態を高い時間精度で可視化することができる。 2 点目は、様々な優れた蛍光特性を有する色素化合物をプローブに導入できることである。このことは、蛍光タンパク質では達成されていない 700 nm 以上の近赤外蛍光を発するプローブや蛍光タンパク質より明るく光退色に強い色素を組み込んだプローブの開発が可能であることを意味している。 3 点目は、比較的小さなサイズのタグタンパク質も利用できることである。タグタンパク質が大きいからといって、必ずしも標的タンパク質の機能に悪影響を与えるわけではないが、一般的に小さなタグタンパク質の方が好まれる。
一方、既存の標識技術では、ほとんどの場合、常時蛍光を放つタイプのプローブを利用していることが問題であるといえる。なぜならば、プローブを細胞に添加しただけでは、多くの場合、タグタンパク質を標識したプローブと遊離プローブの蛍光を区別することができないためである。このことは、細胞を洗浄し遊離プローブを除去することで解決されると考えられる。しかしながら、プローブの構造によっては、洗浄除去に時間がかかる場合や、完全に除去することができないことがあり、迅速なイメージングに支障をきたし S/N 比の低下につながるということが問題となる。そこで、この問題の解決策として、遊離の状態では非蛍光性で、タグタンパク質と結合すると蛍光強度が上昇する「発蛍光プローブ」を用いることで、洗浄操作を行うことなく、迅速にタンパク質をイメージングすることができると期待される(図 1)。発蛍光プローブの先駆けとしては、前述の Tetracysteine tag を用いた手法で用いられる蛍光標識プローブを挙げることができる。しかしながら、過去の報告 6) 及びメーカーのプロトコールによると、遊離プローブは非蛍光性であるものの、プローブの細胞内成分への非特異結合に伴いバックグラウンド蛍光が上昇するため、やはり洗浄作業が必要となる。したがって、洗浄操作を行うことなく迅速かつ高い S/N 比でタンパク質をイメージングすることのできる新しい発蛍光プローブの開発が期待されていた。

3. 発蛍光プローブと Photoactive yellow protein(PYP)タグとによるタンパク質標識技術の開発
著者らは、以上の問題を解決するために、新しいタグタンパク質と発蛍光プローブを利用したタンパク質標識技術の開発に取り組んだ。まず、タグタンパク質を次の 3 つの基準で決定した。(1)動物以外の生物由来のタンパク質の中から探索する。これは、動物細胞で発現するタンパク質を選ぶと、標識反応を行うとき、標的タンパク質以外にその内在性タンパク質が標識されてしまうためである。(2)動物細胞中の小分子と結合することがないことに加え、酵素反応を引き起こすことのないものとする。これは、タグタンパク質の標識反応において細胞内小分子とプローブが競合しないようにするとともに、細胞内代謝に与える影響を最小限にするためである。(3)タンパク質のサイズができる限り小さいものとする。以上の条件のもと、探し出したタンパク質は紅色硫黄細菌 Halorhodospira halophila 由来 PYP であった。
PYP タグは 125 アミノ酸(14kDa)からなる小タンパク質である 7) 。その大きさは、蛍光タンパク質(238 アミノ酸 : 27kDa)の約半分であり、タグタンパク質として魅力的なサイズである。 PYP タグのリガンドとなる化合物は、天然補因子である 4- ヒドロキシ桂皮酸チオエステル誘導体であり、 PYP タグの Cys69 とチオエステル交換反応により結合することが知られている 8) 。さらに、 PYP タグは 7- ヒドロキシクマリン-3- カルボン酸チオエステル誘導体とも結合することが報告されており 9) 、このことは、発蛍光プローブを設計するうえで極めて重要な点である。以前の報告から、クマリンとフルオレセインの誘導体を柔軟なリンカーで連結すると会合消光を引き起こすことが分かっている 10) 。このことから、 PYP タグリガンドのクマリン誘導体とフルオレセインをエチレングリコールリンカーでつないだプローブは、遊離の状態では消光すると考えた。また、 PYP タグとクマリン部位が結合するとフルオレセインとの会合が解消され、蛍光強度が上昇すると仮説を立てた(図 2)。

そこで、この設計指針に基づいて、クマリン誘導体にフルオレセインをつないだ構造を有するプローブ(FCTP)を合成した。クマリン環へのリンカーの導入位置は、タンパク質の立体構造を考慮に入れてクマリンの 5 位とした。
まず、FCTP と精製した PYP タグを反応させ SDS-PAGE(変性ゲルを使用)で解析したところ、タンパク質を示す位置に蛍光バンドが観測されたことから、FCTP と PYP タグは共有結合することが示された。また、細胞溶解液中で FCTP と PYP タグを反応させ、SDS-PAGE で解析したところ、 PYP タグの分子量を示す位置に単一の蛍光バンドが確認できたことから、夾雑タンパク質が存在する条件においても、FCTP と PYP タグは特異的に結合することが示された。
次に、 PYP タグ非存在下で、FCTP の蛍光が消光し、 PYP タグとの結合によって蛍光強度が上昇するかを検討した。遊離状態の FCTP の蛍光スペクトルを測定したところ、蛍光強度は極めて低いことが示された。一方、FCTP と PYP タグを 24 時間反応させると、その蛍光強度は約 20 倍上昇することが明らかとなった。以上の結果から、FCTP は PYP タグを特異的に標識する発蛍光プローブであることが示された 11) 。
4. 発蛍光プローブによるタンパク質の無洗浄生細胞イメージング
FCTP は、OFF/ON 蛍光スイッチを持つ分子であるものの、PYP タグに対する結合速度が極めて遅く(50% 結合時間 : 470 分以上)、実用的な時間で洗浄操作なしのイメージング実験を行うには、更なる結合速度の向上が必要であった。そこで、結合速度を改良すべく第二世代となる発蛍光プローブの開発に乗り出した。
FCTP と PYP タグの結合速度が遅い理由として考えられたことは、リガンド部位であるクマリンと色素部位であるフルオレセインの会合構造が、 PYP タグとの結合において立体障害を引き起こしていることであった。このため、リガンドと色素が会合しない分子構造を有する発蛍光プローブの設計が必要であると考えた。まず、 PYP タグとプローブが結合する際に起こる脱離反応に着目した。結合反応において、チオエステル交換反応によりプローブからチオール化合物が脱離するが、このチオール化合物に色素会合のための消光基を組み込むことで、発蛍光標識と反応速度の向上が同時に達成されると考えた。重要な点は、消光基としてニトロベンゼンを選択したことである。ニトロベンゼンは、種々の蛍光色素との動的・静的消光を引き起こすために、プローブの蛍光色素を変更した場合でも、消光基として汎用的に用いることができると期待される 12) 。分子設計のもう一つのポイントは、リガンド部位をクマリンから桂皮酸に変更することで、蛍光色素とリガンドのπ-πスタッキングを抑制し、反応速度を向上させようとしたことである。このようにして、蛍光色素は、遊離状態ではリガンド部位ではなくニトロベンゼンと会合・消光し、結合反応に伴いニトロベンゼンが脱離し、蛍光色素の蛍光強度が上昇することを期待した。これらの消光基及びリガンドとフルオレセインを色素として組み込んだプローブを FCANB と名付け、合成した(図 3)。

標識反応を行い SDS-PAGE で解析したところ、FCANB と PYP タグが共有結合していることが示され、細胞溶解液中においても、FCANB と PYP は特異的に結合することが明らかとなった。さらに、蛍光スペクトルを測定したところ、FCANB は遊離状態では消光し、 PYP タグとの結合により蛍光強度を 15 倍上昇させた。これらの結果から、FCANB は PYP タグの特異的な発蛍光プローブであることが示された。
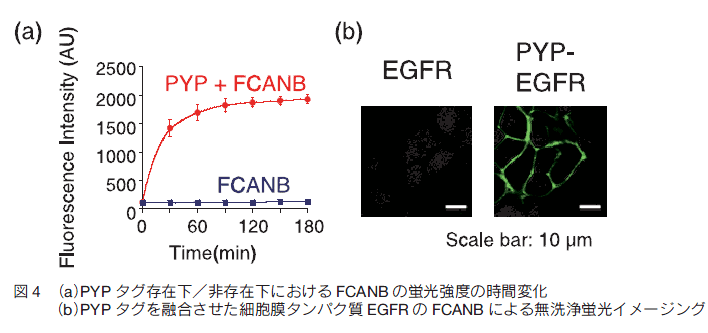
実際に、プローブと PYP タグの反応速度が向上しているかを検討するために、二次反応速度定数 k2 を決定した。その結果、FCANB の k2 は 125 (M-1s-1)であり、FCTP( k2 = 1.11 (M-1s-1))に比べ 110 倍反応速度が向上し、期待したとおりのプローブの改良ができたといえる(図 4a)。
次に、FCANB を用いて、生細胞に発現させた PYP タグ融合タンパク質を洗浄操作なしでイメージングできるかを検討した。 PYP タグには、膜タンパク質の一種である EGFR (上皮成長因子受容体)を融合させ細胞膜上に発現させた。プローブをその発現細胞に添加し、細胞洗浄することなくそのまま共焦点蛍光顕微鏡観察を行った。その結果、細胞膜上から強い蛍光シグナルが観測された(図 4b)。一方、細胞内や培地からはほとんど蛍光が観測されなかった。このことから、生細胞においても特異的に標識反応が起こることが示された。以上により、プローブの反応速度を大きく向上させた発蛍光プローブを用いることで、生細胞膜上の PYP タグ融合タンパク質を洗浄することなく高感度に観測することに成功したといえる 13) 。
5. 環境応答性発蛍光プローブによる生細胞内蛋白質イメージングへの応用
FCANB は細胞膜非透過性であり、細胞内タンパク質を標識することはできなかった。そこで、細胞膜透過性であり細胞内タンパク質を無洗浄で迅速にイメージングすることができるプローブの開発が新たな課題となった。
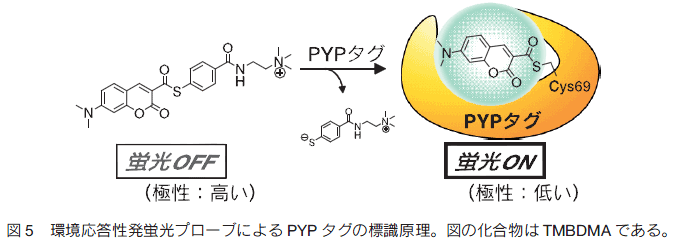
開発にあたり注目したのは、PYP タグのリガンドの分子構造である。現在までに報告されているリガンド分子は、4- ヒドロキシ桂皮酸、4- ジメチルアミノ桂皮酸及び 7- ヒドロキシクマリンの誘導体などである。これらの分子構造の類似性から、7- ジメチルアミノクマリン(DMAC)誘導体も PYP タグに結合すると予想した。この分子に着目したのは、 DMAC は環境応答性蛍光色素であり、極性の高い溶媒中では蛍光強度を低下させ、極性の低い溶媒中では蛍光強度を上昇させる性質を持っているためである 14) 。この性質を応用することで、 DMAC 誘導体を PYP タグの発蛍光プローブとして利用できるのではないかと考えた(図 5)。プローブは、遊離状態では極性の高い水中にあるため蛍光強度が低下し、標識反応により PYP タグの疎水性ポケットにはまり込むために蛍光強度が上昇すると予想した。 DMAC の水溶性向上のため、カチオニックなトリメチルアミノ基またはアニオニックなカルボン酸を導入したプローブ TMBDMA もしくは CMBDMA を設計・合成した。
まず、SDS-PAGE による解析により、両プローブとも共有結合により PYP タグと結合し、細胞溶解液中においてもプローブは PYP タグと特異的に結合することが示された。蛍光スペクトルを測定したところ、プローブの蛍光強度は、遊離状態では低く、PYP タグとの反応により大きく上昇した(TMBDMA : 22 倍、CMBDMA : 16 倍)。このことから、両プローブともに PYP タグをラベル化する発蛍光プローブであることが示された。更に、二次反応速度定数を決定したところ、TMBDMA は k2= 3,950(M-1s-1)、CMBDMA は k2 = 126(M-1s-1)であり、TMBDMA に関しては、 FCANB に比べ反応速度の大幅な向上(約 32 倍)がみられた。
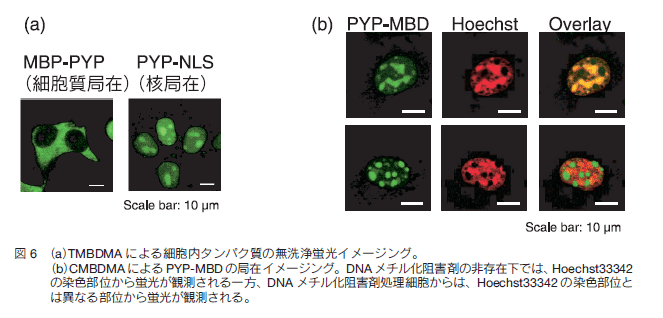
次に、これらのプローブを用いて、生細胞内に発現させた PYP タグを標識しイメージングできるかを検討した。まず、マルトース結合タンパク質 MBP と PYP タグの融合遺伝子 MBP-PYP と、PYP タグと核局在化シグナルの融合遺伝子 PYP-NLS を細胞に導入し発現させ、TMBDMA を添加し洗浄操作を行うことなく共焦点蛍光顕微鏡で観察を行った(図 6a)。その結果、MBP-PYP 発現細胞では主に細胞質から、PYP-NLS 発現細胞では核から蛍光が観測された。一方、非発現細胞からは蛍光は観測されなかった。同様の結果が CMBDMA に関しても得られた。以上の結果から、TMBDMA 及び CMBDMA は細胞膜を透過し、細胞内タンパク質を標識できることが示された。また、これらのプローブを用いることで、洗浄操作無しで生細胞内タンパク質をイメージングすることに成功した。さらに、PYP-NLS 発現細胞に TMBDMA を添加し経時的に蛍光画像を撮影したところ、約 6 分で核内の蛍光が飽和することが分かった。このことから、TMBDMA を用いることにより、極めて短い時間で細胞内のタンパク質をイメージングできることが示された。
最後に、著者らは、PYP タグを用いた標識技術を細胞核内の DNA メチル化のイメージングに応用した。DNA メチル化は、遺伝子発現を制御する重要な化学修飾であり、その修飾反応は DNA メチル転移酵素により触媒され、CpG 配列を持つシトシンの 5 位がメチル化されることが知られている 15) 。まず、メチル CpG に結合するタンパク質である MBD1(MethylCpG-binding domain 1)と PYP タグの融合遺伝子 PYP-MBD を細胞に導入し発現させた。 TMBDMA または CMBDMA を添加し洗浄操作を行わずにイメージングを行ったところ、核内からドット状の蛍光が複数観測され(図 6b)、また、その蛍光は、Hoechst の蛍光と局在が重なった。一般に、Hoechst は、核内のヘテロクロマチンと呼ばれる DNA メチル化の亢進した領域を染色することが知られていることから 16) 、PYP-MBD は、ヘテロクロマチン領域に局在化していることが示唆された。更に、DNAメチル化阻害剤である 5-AzadC を添加し、イメージング実験を行ったところ、PYP-MBD に結合したプローブの蛍光は、Hoechst 染色部位とは異なる位置から観測された。このことは、DNA メチル化阻害剤非存在下では、PYP-MBD が DNA メチル化領域に結合しており、DNA メチル化の阻害に伴う DNA のメチル化レベルの低下により、DNA から解離し局在を変化させたと考えられた。このように、本技術は、DNA メチル化解析に応用することができ、DNA メチル化阻害剤の生細胞評価ツールとしても利用できると期待される 17) 。
6.おわりに
本研究では、PYP タグという新しいタグタンパク質を発掘し、PYP タグを標識することのできる発蛍光プローブを化学の原理に基づき設計・開発した。蛍光色素の分子内会合消光や環境応答性を利用した発蛍光スイッチを活用することで、細胞膜上や細胞内のタンパク質を迅速かつ高い S/N 比でイメージングする技術を開発した。タグタンパク質をラベル化する発蛍光プローブは、いくつかのグループで開発されているが、多くの場合、生細胞内のタンパク質を洗浄操作無しで特異的に検出するのに数十分から数時間を要する。これに対し、開発したプローブの中でも TMBDMA は、数分以内という極めて短い時間での実験を可能にした。このように、迅速に高い S/N 比で生細胞内タンパク質をイメージングできることは、本技術の大きなアドバンテージである。また、PYP タグのサイズが小さいということも、タンパク質の生細胞イメージングにとって理想的な特長といえる。今後は、イメージングの更なる高速化・高感度化に取り組むとともに、蛍光以外の機能も併せ持つ分子をプローブに導入することで、生命科学を探求する新しい化学ツールを創製していきたい。
| 著者プロフィール | |
| 氏名 | 堀 雄一郎(Yuichiro Hori) |
|---|---|
| 所属 | 大阪大学大学院工学研究科、大阪大学免疫学フロンティア研究センター、JST さきがけ |
| 略歴 | 1999 年 京都大学薬学部卒業 2001 年 京都大学大学院薬学研究科修士課程修了 2004 年 京都大学大学院薬学研究科博士課程修了 2004 年 米国ロックフェラー大学博士研究員(Tom W. Muir 研究室) 2006 年 大阪大学大学院工学研究科 助教(現職) 2012 年 JST さきがけ研究者(兼任) 2013 年 大阪大学免疫学フロンティア研究センター 助教(兼任) |
| 出身学校 | 京都大学大学院薬学研究科 |
| 学位 | 博士(薬学) |
| 現在の専門 | ケミカルバイオロジー、タンパク質・ペプチド化学 |
| 氏名 | 菊地 和也(Kazuya Kikuchi) |
|---|---|
| 所属 | 大阪大学大学院工学研究科、大阪大学免疫学フロンティア研究センター |
| 略歴 | 1988 年 東京大学薬学部卒業 1990 年 東京大学大学院薬学系研究科修士課程修了 1994 年 東京大学大学院薬学系研究科博士課程修了 1994 年 米国カリフォルニア大学サンディエゴ校 博士研究員(Roger Y. Tsien 研究室) 1995 年 米国スクリプス研究所 リサーチアソシエイト(Donald Hilvert 研究室) 1997 年 東京大学大学院薬学系研究科 助手 2000 年 東京大学大学院薬学系研究科 助教授 2001 年 JST さきがけ研究者 2006 年 大阪大学大学院工学研究科 教授(現職) 2009 年 大阪大学大学免疫学フロンティア研究センター 主任研究者(兼任) |
| 出身学校 | 東京大学大学院薬学系研究科 |
| 学位 | 博士(薬学) |
| 現在の専門 | ケミカルバイオロジー |
| Copyright(c) 1996-2014 DOJINDO LABORATORIES, ALL Rights Reserved. |